Termite mounds possess various architectural designs. For example, in Macrotermes bellicosus, the structures vary depending on their location (Collins 1979). When situated in savanna habitats, colony mounds are dense and possess numerous ridges and thin walls. In the cooler and more temperature-stable gallery forest, mounds are shaped dome-like with thicker walls to reduce heat loss. A mound structure may help regulate warm temperatures, but sufficient gas exchange is necessary for survival. There is an apparent trade-off between gas exchange and thermoregulation (Korb & Linsenmair 1999). In areas of sub-optimal temperatures, increased thermal insulation through structural additions resulted in elevated levels of CO2. High CO2 levels reduce the metabolic productivity of the symbiotic fungi that aid termites in breakdown of cellulose.

Macrotermes within a basidiomycete fungal comb
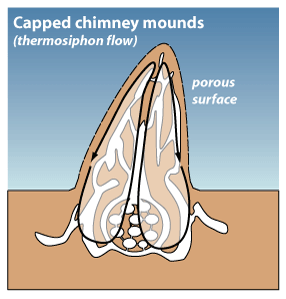 Martin Lüscher's thermosiphon model
Martin Lüscher's thermosiphon model
Evaporative cooling also aids in thermoregulation. If the walls of a termite mound are kept moist, water vapour is able to diffuse though pores of the soil. Termite colonies are capable of maintaining humidity by transporting water contained within earth in and out of the mound (Turner, 2001). Species of the subfamily Macrotermitinae share an endosymbiotic relationship with basidiomycete fungi, their metabolism tripling or quadrupling by the fungus activity assisting in digestion of cellulose (Korb & Linsenmair 2000; Abe et al. 2000). Optimal growth of the fungi requires constant temperatures of 30˚C, high humidity, and low concentrations of CO2 . The structures of the fungi are additionally able to absorb humidity and release water in order to stabilize humidity and prevent other the competition between other fungal species that prefer certain humidity levels.
Currently, the mechanism of gas exchange between termite colonies and the surrounding air remains largely unknown. Since the late 1950s, the “thermosiphon ventilation” theory of entomologist Martin Lüscher had been widely accepted (Turner 2001). He proposed that termite mounds functioned similarly to a circulatory system in capped-chimney mound termites. He hypothesised that the metabolic heat produced by the colony humidifies the air and reduces its density, rising into the mound (Lüscher, 1956, 1961). This forces air below into the nest tunnels below the mound surface, which are sites for loss of excess heat and gas exchange. The air becomes denser and sinks down to the nest, in which the process repeats the circuit again. A higher metabolism rate would result in a greater buoyant force, therefore leading to a stronger circulation. Lüscher’s model suggests metabolic-induced convection is the driving force of airflow between the mound and nest.
Studies by J. Scott Turner (2001) on Macrotermes michealseni show the metabolic heat generated is not substantial enough in order to drive gas exchange alone. The metabolic heat produced over an hour is estimated to equal 200-400 kilojoules, enough to warm nest temperature by only a tenth of a degree.
Korb and Linsenmair (1999) also sought to test Lüscher’s thermosiphon ventilation mechanism, however it was not supported by their results. Instead, a temperature gradient is formed between air channels that are warmed during the day and the cooler upper section of the mound. This results in convection currents, air rising from the air channels to the top of the mound and exchanging CO2. Additionally, by comparing temperatures of inhabited and uninhabited mounds, they found mound structure resulted in a relatively constant nest temperature (Korb & Linsenmair, 2000). Only large colonies with mounds above 2 metres were able to reach optimal nest temperatures of 30˚C. A larger colony with a more active metabolism and possibly an increased ability to make structural alterations is related to high nest temperatures.
It appears that nest temperature and soil temperature are linked, and the embedding of the nest within the soil is able to considerably damp temperature fluctuations (Turner 2001). Despite this, it was suggested that metabolic-induced buoyant forces might still play a role in homeostasis of termite colonies in conjunction with wind energy interaction. Turner’s own model proposes that gas exchange is driven by the combination of metabolic-induced buoyancy, wind energy, and the interaction between the mound and nest structure. Instead of Lüscher’s circulatory model, Turner describes the ventilation as ‘tidal’ in nature, depending on the fluctuation of wind.
Currently, the mechanism of gas exchange between termite colonies and the surrounding air remains largely unknown. Since the late 1950s, the “thermosiphon ventilation” theory of entomologist Martin Lüscher had been widely accepted (Turner 2001). He proposed that termite mounds functioned similarly to a circulatory system in capped-chimney mound termites. He hypothesised that the metabolic heat produced by the colony humidifies the air and reduces its density, rising into the mound (Lüscher, 1956, 1961). This forces air below into the nest tunnels below the mound surface, which are sites for loss of excess heat and gas exchange. The air becomes denser and sinks down to the nest, in which the process repeats the circuit again. A higher metabolism rate would result in a greater buoyant force, therefore leading to a stronger circulation. Lüscher’s model suggests metabolic-induced convection is the driving force of airflow between the mound and nest.
Studies by J. Scott Turner (2001) on Macrotermes michealseni show the metabolic heat generated is not substantial enough in order to drive gas exchange alone. The metabolic heat produced over an hour is estimated to equal 200-400 kilojoules, enough to warm nest temperature by only a tenth of a degree.
Korb and Linsenmair (1999) also sought to test Lüscher’s thermosiphon ventilation mechanism, however it was not supported by their results. Instead, a temperature gradient is formed between air channels that are warmed during the day and the cooler upper section of the mound. This results in convection currents, air rising from the air channels to the top of the mound and exchanging CO2. Additionally, by comparing temperatures of inhabited and uninhabited mounds, they found mound structure resulted in a relatively constant nest temperature (Korb & Linsenmair, 2000). Only large colonies with mounds above 2 metres were able to reach optimal nest temperatures of 30˚C. A larger colony with a more active metabolism and possibly an increased ability to make structural alterations is related to high nest temperatures.
It appears that nest temperature and soil temperature are linked, and the embedding of the nest within the soil is able to considerably damp temperature fluctuations (Turner 2001). Despite this, it was suggested that metabolic-induced buoyant forces might still play a role in homeostasis of termite colonies in conjunction with wind energy interaction. Turner’s own model proposes that gas exchange is driven by the combination of metabolic-induced buoyancy, wind energy, and the interaction between the mound and nest structure. Instead of Lüscher’s circulatory model, Turner describes the ventilation as ‘tidal’ in nature, depending on the fluctuation of wind.
